a figure from the essay
Fossils, Genes and the Evolution of Animal Limbs
by Neil Shubin, Clif Tabin and Sean Carroll
Figure 2 (from page 104 of Shaking the Tree: Readings from Nature in the History of Life edited by Henry Gee)
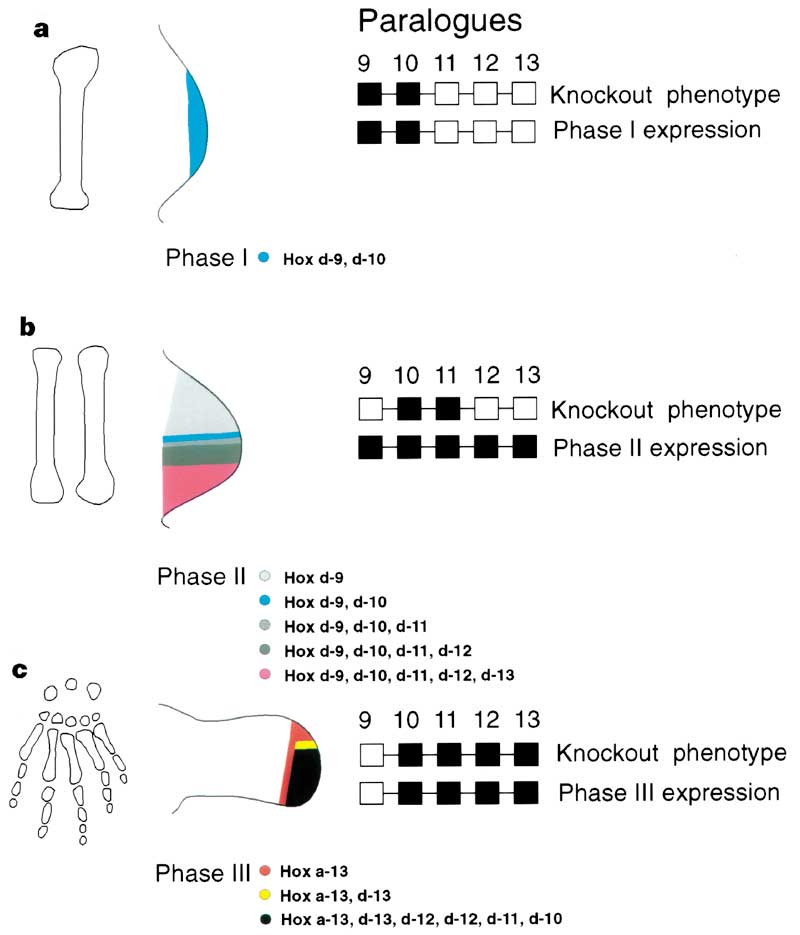
Stylopod, zeugopod and autopod: patterning the limb. The tetrapod limb consists of three distinct compartments: a, the stylopod (upper arm and thigh); b, zeugopod (lower arm and calf); and c, autopod (hand and foot). This subdivision of the limb is supported by phylogenetic comparison,4 analysis of gene expression,7 and experimental manipulation.14,36-41,109-111 There is a broad correlation between the position of a compartment and its evolutionary history.4 The stylopod (a) is the most ancient (possibly of late Silurian origins) whereas the zeugopod and autopod are the most recent (being first encountered in Devonian sarcopterygians). This same order of appearance of the three limb segments is recapitulated during development. Early removal of the apical ectodermal ridge (AER) results in a limb with only a stylopod; the zeugopod and autopod are produced after successively later surgeries.112,113 The Abd-B-related genes of the HoxD cluster are expressed in a complex, dynamic pattern encompassing at least three distinct,7 independently regulated phases.24,26 In the first phase (a; phase I), two of these genes (HoxD-9 and HoxD-10) are expressed across the entire limb bud.7 This expression correlates with the time that the stylopod is specified.7,112 Subsequently, a second phase of expression (b; phase II) is initiated in response to the secreted factor, Sonic hedgehog. Here, Hox genes are expressed in a nested set centered around the Sonic-expressing cells, with HoxD-13 being expressed in the most restricted domain, and HoxD-12 and HoxD-11 each encompassing a broader domain.7 This pattern of expression coincides with the time of specification of the zeugopod and takes place in cells fated to form this segment. Finally, a third phase of expression (c; phase III) is initiated later during limb development, when these Hox genes are all expressed across the majority of the distal portion of the limb bud.7 During this phase, the expression of Hox genes still appears to be a consequence of the Sonic hedgehog signal, but the relative responsiveness of the different genes has changed so that HoxD-13 now has the broadest expression domain and HoxD-12 and HoxD-11 are nested within it.7 The phase III expression (c) patterns occur in the presumptive autopod at the time that segment is specified. The combination of the change in relative size of Hox expression domains with a phenomenon known as "posterior prevalence" (the general rule that more 5' genes in the Hox cluster are phenotypically dominant) results in different Hox genes playing pre-eminent roles in different limb segments: for example, HoxD-9 during phase I in the stylopod (a), HoxD-11 during phase II in the zeugopod (b), and HoxD-13 during phase III in the autopod (c). The expression of the dominant Hox genes in each phase is essential for the formation of the bones in each segment, as seen in their knockout phenotypes. The knockout phenotype of a gene consists of alterations in the pattern and shape of skeletal elements. The location of these modifications depends on the position of the gene within the cluster. For example, mice engineered to be deficient in both HoxD-11 and HoxA-11 (the paralogous gene of the HoxA cluster) form limbs that are essentially missing the zeugopod14 (b). Phenotypes of Hox D-9-deficient mice, in contrast, are specific to the stylopod109 (a), whereas Hox D-13-deficient mice primarily have defects in the autopod39 (as do mice engineered to be deficient of Hox A-13 paralogues;110,111 c). Knockout data are derived from refs. 14, 36-41 and 109-111.
Back to the list of color illustrations.
About the book: Shaking the Tree: Readings from Nature in the History of Life edited by Henry Gee, published by the University of Chicago Press.